Practica 3° Observación de liquen
Los líquenes son asociaciones simbióticas entre un alga (fotobionte) y un hongo (micobionte) de cuya interacción se origina un talo estable, con estructura y fisiología específicas. Las algas encontradas pueden ser clorofitas o cianofitas y los hongos que intervienen en la asociación son Ascomycetes, Basidiomycetes o Phicomycetes (Alexopoulos y Mims 1985). Poseen características particulares como carecer de raíz y de sistemas de conducción, no poseer estructuras selectivas o protectoras del medio externo como cutículas o epidermis, por lo que se vuelven vulnerables a variaciones ambientales. Son organismos autótrofos, su crecimiento es muy lento, poseen una amplia distribución desde los polos al ecuador, y crecen en las superficies de los más diversos sustratos inertes u orgánicos. Los líquenes poseen órganos apendiculares cuyas formaciones son producidas por el hongo; al proyectarse desde la cara inferior, sirven para sujetar el talo al sustrato y pueden actuar reteniendo agua (Izco 2000). Los líquenes son importantes ya que pueden ser usados como bioindicadores, absorben el dióxido de azufre, reducen la contaminación, entre otros. En este trabajo se realiza una recopilación, en lo que se refiere a las estructuras, basado fundamentalmente en la propia observación y experiencia. Se realizaron diferentes cortes de las formas biológicas ó biotipos (Folioso, Gelatinoso, Fruticuloso y Ascocarpo) para observar la morfología y anatomía (HOMOMEROS y HETEROMEROS) del talo.
OBJETIVOS
Objetivo general
Reconocer los diferentes tipos y estructuras que hacen distinguibles a los líquenes delos líquenes y de las demás plantas no vasculares.
Objetivos específicos
- Observar las estructuras de los líquenes que se ven a través del microscopio después de un corte especifico
- Identificar el liquen observado por sus estructuras
- Sellar placas con gelatina de glicerina con muestras de líquenes para su respectiva presentación
MARCO TEÓRICO
Los líquenes comenzaron a ser considerados como integrantes del mundo de los hongos a partir de 1866 (HALE, 1967). Es desde esta fecha, cuando se introducen ca racteres, microscópicos, químicos y aquellos que hacen referencia a la estructura de los ascocarpos. Son por tanto los caracteres referidos al componente fúngico y aquellos frutos de la simbiosis, como los químicos, los que han venido delimitando el estado en el que actualmente se encuentra la sistemática de los líquenes.
Por tanto, dada la poca importancia concedida al ficobionte a nivel de división, subdivisión y orden (POELT, 1973 a) y desde el punto de vista de la actual sistemática, los ascolíquenes se incluyen dentro de la división Eumycota, en la subdivisión Ascomycotina (WEBSTER, 1980; POELT & VEZDA, 198 1 ), obviando los basidiolíquenes. La problemática vuelve a aparecer a nivel de clase, mientras algunos autores incluyen la mayoría de los ascolíquenes dentro de la clase Discomycetes (WEBSTER, 1980), otros (HENSSEN & JAHNS, 1974; POELT & VEZDA, 1981) no se pronuncian por el momento, clasificando los hongos liquenizados en órdenes y sin incluir éstos en clases.
Los líquenes durante períodos desfavorables pierden, para sobrevivir, su capacidad de crecimiento, permaneciendo en lo que POELT (1973 a) denominó como «estados especiales de resistencia». Además, como organismos longevos que son, están sujetos durante largos años a los efectos de unos factores medio-ambientales extremos y, en muchos casos, hostiles. Esto podría inducir a considerables modificaciones morfológicas que produjeran una secuencia de variaciones fenotípicas tan grande, sobre todo en géneros de talos crustáceos, que lleguen a expresarse genotípicamente diferenciando especies o incluso grupos (POELT, 1965). Por el contrario, existen fenótipos de determinadas especies que, cuando se desarrollan en hábitats diferentes, pueden parecer tan distintos que solo tras un detallado análisis son reconocidos como pertenecientes al mismo taxón específico e incluso a la misma población.
La vida en simbiosis de algunos hongos superiores con un alga es una especialización encaminada a la obtención de sustancias orgánicas, y se da en grupos taxonómicos de hongos muy diferentes y no demasiado emparentados filogenéticamente (HENSSEN & JAHNS, 1974). Para la mayoría de estos hongos la simbiosis es obligatoria ya que en la naturaleza no se han encontrado, hasta ahora, separados del alga o, en todo caso, fructificados; en el laboratorio, al cultivarlos, en ausencia del ficobionte no fructifican y degeneran, de todas formas el número de experimentos de este tipo y el número de taxones usados ha sido muy reducido. El alga, en cambio, no es estrictamente específica para cada micobionte ya que diferentes hongos liquenizados pueden presentar el mismo ficobionte y es capaz de realizar su ciclo vital completo fuera de la simbiosis, tanto en la naturaleza como al cultivarlas aisladamente (AHMADJIAN, 1 973). En lo que se refiere a biomasa, el micobionte es el componente dominante en la simbiosis y es además el responsable de las complicadas estructuras talinas, aunque la influencia del alga es vital en orden a los resultados morfológicos finales. Los modelos estructurales vegetativos alcanzan, en los líquenes, un alto nivel de diferenciación paralelo, en muchos casos, al de algunas estructuras de plantas superiores (JAHNS, 1973). El ficobionte, en un reducido número de casos, puede llegar a dominar en biomasa, en esta ocasión siempre se trata de algas verde-azuladas (Cyanophytu). En la simbiosis liquénica, el hongo es el único que realiza procesos de reproducción sexual y en el caso de ascolíquenes ésta es bastante semejante a la del resto de los ascomicetes no liquenizados. No obstante, solo se originan dos tipos de estructuras reproductoras: por un lado los filamentos microsporógenos (=conidióforos o fialidios) que producen mitoexósporas (=picnidiósporas, conidiósporas o esparmacios) que pueden ejercer la función de micrósporas (HENSSEN, 198 1) y se originan en unos mitoesporocistes llamados Picnidios; y por otro, filamentos procedentes del ascogonio (ascogoniales) originando los ascos, que contienen a las ascósporas, en el interior de una fructificación llamada ascocarpo (LETROUIT-GALINOU, 1973). De todo lo anteriormente expuesto se puede deducir que es el hongo liquenizado el que cobra mayor importancia en los distintos aspectos del estudio de los líquenes.
Estructura del talo
El proceso de liquenización, puede encontrarse en diferentes grados de estabilización, en función de lo cual se originarán determinadas y específicas estructuras en los distintos grupos de líquenes. Pero en ciertos casos en que esta relación no está definitivamente estabilizada, la anatomía y morfología de estos líquenes recuerdan o bien a las del alga o bien a las del hongo en estado de vida libre. Hasta ahora no se ha formulado una teoría, lo suficientemente fundamentada y generalizable, que relacione la estructura del talo con las posibles líneas evolutivas y origen mono o polifilético de los líquenes. Aunque de forma intuitiva todos los autores coinciden en aceptar el polifiletismo de este grupo de organismos (AINSWORTH, 1971; POELT & VEZDA, 1981).
De una manera un tanto artificial, pero cómoda, se han distinguido tradicionalmente (OZENDA & CLAUZADE, 1970; JAHNS, 1973; HENSSEN & JAHNS, 1974) como tipos fundamentales de estructuras los siguientes:
ESTRUCTURA HOMOMERG ES característica de los talos gelatinosos, de organización más simple e indiferenciados, en los que el ficobionte (p.e. Nostoc) está irregularmente distribuido en cenobios filamentosos que desarrollan una matriz gelatinosa donde crece el micobionte. Esta estructura se encuentra relegada, en su sentido más estricto, al género Collema. Sin embargo hay que resaltar que, dentro de la misma familia (Collemataceae), se observa una complicación en el género Leptogium, donde aparece un estrato paraplectenquimático corticando al resto del talo, es lo que se denomina ((Estructura Celúlica~ que como señalaba ya DEGELIUS (1954) parece que puede representar el paso a la heteromena (Fig. 3 a y b). 11.
ESTRUCTURA HETEROMERA. ES la más corriente y compleja. El ficobionte queda restringido a una capa particular y delimitada por otras capas diferenciadas en las que no aparecen los gonidios. Normalmente los estratos se suceden horizontalmente (Estructura Heterómera Estratificada), pero una variante viene dada en algunos de los táxones con biótipo fruticuloso, en donde las capas se disponen radialmente alrededor de un eje central (Estructura Heterómera Radiada). Ver Fig. 3 c, d, e y g.

IMAGEN TOMADA DE: (Barreno and Rico, 2018)
Existen otros dos tipos que podemos considerar extremos y que a veces son confundidos con los modelos estructurales básicos y en muchas ocasiones recuerdan al talo de algas y hongos de vida libre. Así ocurre en el género Lepraria, en el que no se aprecia una estructura diferenciada sino pulverulento-granulosa, donde el micelio y los gonidios se encuentran laxamente entremezclados (Estructura Lepraroide). A veces ocurre en talos de otros géneros, probablemente por influencia de factores externos al 1í- quen, P.e. efectos de la contaminación ambiental o falta de luz, ya que cuando estos factores no inciden la estructura no se altera. Otro caso es el de la Estructura Filamentosa, a veces referida a la homó- mera, que presentan algunos líquenes con talos de biótipo fruticuloso de pequeñísimo tamaño y que están formados por filamentos del alga (Scytonema, Stigonema), causante de este tipo morfológico, envueltos por unas pocas hifas del hongo (Fig. 3 f).
Esta estructura se da en un reducido número de táxones incluidos en la familia Lichinaceae. Ya que la gran mayoría de los 1íquenes presentan la estructura más compleja, heterómera, es conveniente detenerse en la descripción de cada una de las capas o estratos que presentan estos talos. De fuera a dentro son:
1I.a. EPICORTEX (=Capa Epinecral). Se trata de un estrato sin células, compuesto por una fina capa de polisacáridos que recubre la parte exterior del talo y se sitúa directamente en contacto con el medio ambiente (HALE1976). Se le supone análoga función que a la cutícula en plantas superiores. Muchos táxones liquénicos no poseen esta capa de protección, sino que las células fuertemente empaquetadas de las hifas corticales son las encargadas de hacer frente al medio. Ninguno de los autores consultados hasta ahora, relaciona a la pruína, que está constituida por gránulos de aspecto escamoso, fruto de la acumulación de carbonatos y oxalatos y que aparecen cubriendo exteriormente a cierto número de talos, con esta capa sino con el córtex, a pesar de no poseer ninguna célula (JAHNS, 1973). (Fig. 3 g).
1I.b. CORTEX. La mayoría de los 1íquenes se encuentran protegidos por una capa cortical de células del hongo, muchas veces diferencialmente pigmentada, siempre recubriendo la parte superior del talo (Córtex Superior) e inmediatamente debajo del epicórtex, si existe. A veces también cubre la cara inferior (Córtex Inferior), como es el caso de algunos talos sin estructura radiada. El grosor de esta capa varía según los táxones y no siempre forma un estrato continuo, sino que puede quedar interrumpido por penetraciones de la capa gonidial u otras estructuras más específicas (hendiduras, pseudocifelas, cilios, etc.). El córtex puede estar formado por cualquier tipo de plecténquima de los descritos con anterioridad. Ocasionalmente dos tejidos diferentes pueden combinarse en esta capa, sobre todo en talos fruticulosos (Coelocaulon, Cetraria, etc.) pudiéndose diferenciar por su distinta orientación (Fig. 3 c). No existe, por ahora, un criterio lo suficientemente claro como para poder definir y denominar los distintos tipos de córtex de los talos liquénicos, ya que se han venido estudiando por simple comparación pero sin tratar de interrelacionarlos o de establecer sus relaciones naturales. No obstante existen un par de términos que sirven para definir extremos de variación de la estructura cortical: «Pseudocórtex» (DEGELIUS, 1954), originariamente descrito para nominar las capa cortical de algunos táxones del género Collema y que está compuesto por células algo engrosadas del ápice de las hifas, en número variable, irregularmente desarrolladas y no constante ni claramente delimitadas con respecto a las hifas del interior del talo. HANNEMANN (1973), describe un «Corticoide» (=Scheinrinde p.p. sensu POELT, 1958) en el que la estructura del córtex está un tanto alterada por la presencia de células algales de la capa gonidial, las células de las hifas del hongo no están bien definidas y se encuentran bastante dispersas. Este último tipo es frecuente en muchos talos crustáceos en los que tanto el córtex como la médula y la capa gonidial están estructuralmente influenciados por el ficobionte y son difíciles de separar entre sí. La diferencia fundamental entre estos dos conceptos viene dada por el tipo de alga constituyente del líquen. En el primer caso se trata del género Nostoc (verde-azulada) y en el otro son cloroficeas (Trebouxia, Trentepohlia, etc.) las que intervienen. No obstante la semejanza de las dos estructuras es manifiesta. (Fig. 3 a).
1I.c. CAPA GONIDIAL (Capa Algina). Aquí es donde se hace patente la dualidad del complejo liquénico. La relación entre alga y hongo varía en función del tipo e intensidad del contacto. Puede ocurrir que el hongo no se encuentre en relación directa con el alga, sino que presione contra las células del ficobionte mediante una formación llamada «Apresorio» (JAHNS, 1973). Otras veces el haustorio penetra a través de la membrana de la célula algal.
Obviamente esta capa queda delimitada por la presencia del alga. Su posición y grosor vana en los distintos géneros, situándose siempre donde la intensidad lumínica es suficiente, que es por lo general, inmediatamente debajo del córtex superior. Puede estar desarrollada desde en capas contínuas y con + igual grosor, hasta en capas irregulares, o en glomérulos aislados a lo largo del talo. La multiplicación del alga, dentro de la capa, se hace por aplanósporas o por simple división mitótica de las células (JAHNS, 1973).
El ficobionte más corriente en los líquenes es unicelular y verde perteneciente al género Trebouxia. HENSSEN & JAHNS (1974) calculan que el 83% de los líquenes de Escandinavia llevan estas algas. Otros géneros de gonidios verdes no son raros, como el filamentoso Trentepohlia o el unicelular Pleurococcus, en talos crustáceos comprendiendo un 9% de la flora. Las algas cianoficeas se integran en el 8% restante de los talos de la flora liquénica escandinava, y se incluyen géneros como Nostoc, Scytonema, Stigonema, etc. propios de talos gelatinosos, filamentosos y foliaceos pero con tonalidades oscuras.
A pesar de que existen claves para la determinación de los géneros del ficobionte que entra a formar parte del talo liquénico (AHMADJIAN, 1967), 10s conocimientos actuales sobre la especiación y constancia del alga son extremadamente limitados y muchos miles de líquenes quedan por ser estudiados.
1I.d. MEDULA. ES normalmente el plecténquima medular el encargado de formar este estrato, aunque en otros casos, y dependiendo de los géneros, puedan intervenir un proso ó un paraplecténquima. Dadas las características del tejido que la forma, posee una gran capacidad de almacenamiento de agua y sustancias liquénicas, además de cristales de oxalato cálcico que se depositan extracelularmente, en la pared de las hifas (JAHNS, 1973). Es preferentemente en la médula donde se acumulan la mayor parte de las sustancias liquénicas y es también generalmente en esta capa donde se originan y desarrollan, en sus primeros estadíos y hasta que salen al exterior, los ascocarpos (OZENDA & CLAUZADE, 1970; LETROUIT - GALINOU, 1966; HENSSEN, 1981) extendiéndose más o menos pronto hacia la capa gonidial y el córtex.
El grosor del estrato vana también según los géneros aunque hay que destacar que suele ser una capa comparativamente más grande que el resto. Está delimitada en la parte superior: por la capa gonidial, con la que está en contacto directo, e incluso con el córtex superior si aquella es discontinua. Por la inferior con: el sustrato, en el caso de los líquenes crustáceos; directamente con el ambiente aéreo, en los fruticulosos sin cortex inferior (p. e. Evernia); con el cortex inferior, si se trata de talos foliáceos que lo posean o, de nuevo con la capa gonidial en el caso de algunos fruticulosos de estructura estratificada y siempre en la radiada. Existen otros dos tipos de estratos (iformaciones?) que, dependiendo de los autores (OZENDA & CLAUZADE, 1970; JAHNS, 1973; HENSSEN & JAHNS, 1974; HALE, 1976) son adjudicados o no a la médula. En algunos líquenes fruticulosos de estructura radiada (Usnea) se distingue en el centro del talo un «Cordón Axial» formado por hifas prosoplectenquimáticas, densas y orientadas longitudinalmente, lo que produce una considerable solidez y flexibilidad al talo en estado húmedo (JAHNS, 1973; MOTYKA, 1936-1938). Otros autores consideran, a este cordón de las usneas, como una parte bien definida de la médula, médula intema (OZENDA & CLAUZADE, 1970), o como una simple continuación de aquélla sin denominación específica (POELT, 1969). En otros grupos este cordón está sustituido por una capa de tejido gelatinoso o esponjoso (Ramalina, Alectoria, Cladonia, etc ...) (Fig. 3 d, y e).
Destacamos, por su abundancia en líquenes crustáceos, una estructura libre de algas llamada «hipotalo» (=protalo, prototalo, p.p.), que consiste en una fina capa de hifas, compuestas por células más o menos irregulares, anastomosadas y con la pared carbonizada. Un factor importante del mayor o menor desarrollo de este estrato lo juega la naturaleza del sustrato (WEBER, 1977), en sustratos duros aparece más desarrollado que en blandos. Se puede situar tanto en el córtex inferior, como entre las areolas del talo, extendiéndose irregularmente sobre el sustrato (gen. Rhizocar pon) o inmediatamente debajo de la médula que por otro lado es su posición más normal. En cuanto a su origen existen distintas opiniones, según los autores (JAHNS, 1973; RUNEMARK, 1956; POELT, 1969; HENSSEN & JAHNS, 1974), pero lo que parece claro es que vana según los táxones, en unos evoluciona a partir de la médula y en otros posiblemente del córtex, en función de sus relaciones con el sustrato y competencia por espacio con otros líquenes de la comunidad. Parece jugar un papel importante en la colonización de rocas y cortezas. (Fig. 3 g).
Formas biológicas (Formas de crecimiento, biótipos)
Fue a principios del siglo XIX cuando se realizaron los primeros ensayos para sistematizar los líquenes, tomando como base las diferentes formas de crecimiento que adoptan estos talófitos, separando las clases en función de los principales biótipos (ACHARIUS, 1798; MASSALONGO, 1855, etc. in POELT, 1973a). Así reconocieron tres formas de líquenes que podían crecer tanto en rocas como árboles o suelos: aquellos que forman una fina costra 2 bien desarrollada y difícilmente separable del sustrato (crustáceos); los que con forma -+ de hoja se encontraban más laxamente unidos al sustrato (foliáceos); y por último aquellos que creciendo + ascendentemente, pueden ramificarse hasta adquirir una forma de pequeño arbusto (fruticulosos). Hasta principios de este siglo se han venido utilizando estos conceptos + modificados, para diferenciar incluso familias (HARMAND, 1905 – 19 13; ZAHLBURCKNER, 1922-1940). Ciertamente, las formas de crecimiento en los talos liquénicos no pueden considerarse como caracteres fundamentales en taxonomía. Por ejemplo, los líquenes foliáceos no forman un grupo taxonómico en el que todas las especies se encuentren estrechamente relacionadas, sino que constituyen simplemente una unidad morfológica o fisiológica. En una misma familia, y a veces en un mismo género, pueden presentarse distintas especies con diferentes biótipos, ya sean crustáceos, foliáceos o fruticulosos. Pero no solo se da el hecho de que dos especies con el mismo biótipo no se encuentren taxonómicamente relacionadas, sino que existen numerosas formas de crecimiento intermedias entre las tres fundamentales. Estas formas biológicas tradicionales no constituyen nada más que puntos en una escala continua de diferenciación de talos, desde primitivos hasta altamente estructurados (HALE, 1967). Algunos autores (REINKE, 1894-1896 in JAHNS, 1973) consideran a los líquenes fruticulosos como los que ocupan los puntos más altos en el desarrollo de las diversas líneas paralelas de la evolución de estos talófilos. Esta teoría, aunque sugestiva, no se corresponde con la realidad ya que la estructura de los talos foliáceos no es menos complicada que la de los fruticulosos, por el contrario algunas estructuras altamente diferenciadas solo se encuentran en táxones foliáceos, como las cifelas en el género Sticta. No ocurre así con los líquenes crustáceos, que pueden ser considerados como más primitivos o, en todo caso, secundariamente derivados (POELT, 1973a). Como consecuencia, se puede decir que, estas formas de crecimiento se caracterizan por poseer una particular combinación de sus distintas capas (cortical, medular, gonidial), y por un diferente desarrollo de sus estructuras vegetativas. La apariencia externa y la estructura interna son en todos los casos interdependientes. A pesar de todas las objeciones mencionadas, creemos conveniente una separación de los líquenes en biótipos. La forma de crecimiento es la característica más obvia para distinguir especies de líquenes, además de que suelen ser el punto de partida más común de las claves artificiales de determinación. Como antes se comentaba, existe una gran variabilidad entre las formas clásicas de crecimiento. Atendiendo a ésto y según los diferentes autores (KLEMENT, 1955; HALE, 1967; JAHNS, 1973; HENSSEN & JAHNS, 1974), los biótipos fundamentales, con sus características y variaciones más constantes dentro de cada grupo, que se pueden reconocer son los siguientes:
FOLLÁCEOS: Son laminares, lobulados y con simetría dorsiventral. Presenta talo heterómero. Están parcialmente adheridos al sustrato, por lo que se les puede separar de él sin destruirlos.
FRUTICULOSO: Presentan formas de pequeños arbustos, son erectos o colgantes y con simetría radial, a veces dorsiventral. El tamaño es muy variado: desde menos de 2 cm, pequeños arbustos, hasta los que tienen varios metros de largo y dependiendo de la humedad relativa del aire para su hidratación (aereohigrófilos) (Barreno, 2003). Son heterómeros con simetría radial, macizos o huecos (talo cilíndrico) o heterómeros con simetría dorsiventral (talo aplanado) (Chaparro de Valencia, 2002)
CRUSTACEO: Estos carecen de córtex inferior y están en estrecho contacto con el sustrato, por lo que son difíciles de separarlos de él. Presentan talo de la mayoría de estos líquenes está constituido de pequeñas escamas denominadas areolas.
GEELATINOSO: El ficobionte es el responsable de la forma yconsistencia. El alga es en general verde-azulada. Tiene un talo homomero, con córtex (Leptogium) o sin cortex (Collema). Son rígidos y frágiles cuando están secos. Puden absorber de 20 hasta 30 veces su peso en agua y la coloración en estado húmedo va de verde oliva hasta gamas de gris (Chaparro de Valencia, 2002)
TALOS DIMÓRFICOS: Están constituidos por un talo horizontal (TALO PRIMARIO) adherido al sustrato y otro vertical que lleva los cuerpos fructíferos (TALO SECUNDARIO Ó PODEDO). El talo horizontal puede ser crustáceo (Dibaeis baeomyceas) ó folioso (Cladonia). Algunas veces, en estado adulto, solo se conserva la parte vertical o talo secundario porque el talo primario es fugaz, como en Cladina (Chaparro de Valencia, 2002)
MATERIALES
- Liquenes (Folioso,Gelatinoso, Fructoso, Ascocarpo)
- Agujas de disección
- Cuchilla minora
- Porta y cubreobjeto
- Gotero
METODOLOGÍA
En esta practica se realizaron distintas observaciones a líquenes para ver las estructuras de los diferentes tipos de líquenes utilizamos agujas de disección y cuchillas minoras para tomar un pedazo muy delgado de cada una de los 4 tipos de líquenes que tenían cortes diferentes, el liquen folioso es una corte vertical tomando de su parte superior a su parte inferior muy delgado, en el liquen gelatinoso se repite el mismo proceso que el folioso, el liquen fruticuloso es una corte a través de su tallo en forma muy delgada, para el ascocarpo se toma de un liquen bien formado su parte que sobre sale de este se toma y se corta de forma superior a inferior de forma muy delgada hay que recordar de tener muy bien hidratados estos cortes para mejorar la visión de sus estructuras al tener estos cortes se ponen sobre el respectivo portaobjetos, se observan por el microscopio a 10x en ocaciones para reconocerlos bien se tiene que sellas antes de identificarlos para poder observarlos en 40x ya teniendo esto procedemos a terminar la practica sellando las muestras que se le lograron identificar las estructura
RESULTADOS
Liquen folioso: Tienen una forma o aspecto parecido de lechuga, presenta color verde oscuro. En la imagen se detecta un corte transversal del liquen mencionado, donde se identifica tres colores, los cuales son el verde, el gris y el café; en la verde se evidencia las algas, en la parte café son las capas o epidermis inferiores y superiores, y el color gris es la mezcla del tejido fotosintético y epidermis. Por su organización se determina heterómero.






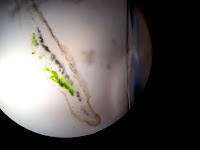

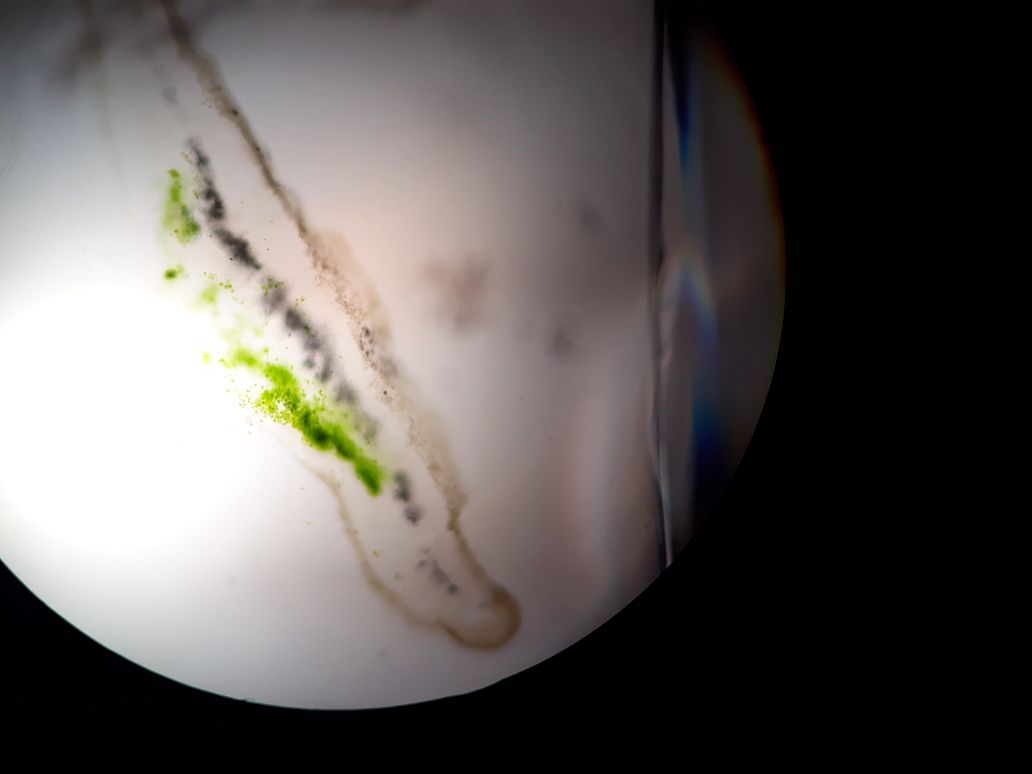
Liquen fruticuloso: Al ser observado el corte transversal de la muestra correspondiente a este liquen se encuentra una simetría que divide en dos planos, dejando ver en el una corteza, capa algal, médula hueca y en algunas ocasiones se logra encontrar a manera de masa compacta solida. Se evidencia una organización de colores los cuales son el verde-amarillo, café y gris. Por su organización se determina heterómero.
Liquen gelatinoso: Talo gelatinoso, verde azulado a negro; lóbulos pequeños a grandes; homómero, sin capas internas distintas, pero con corteza superior e inferior.Se identifica en la imagen un corte transversal, se observa un talo de hifas de forma ovalada que se encuentra en todo el liquen por esto se observa en microscopio y es de color verde-amarilla.
Ascocarpo: Tiene forma de copa con una cobertura no muy fina, se encuentra con un aspecto coloreado. Se realizo un corte transversal. Sacando una tira delgada para poder evidenciar la estructura presente, de una coloración translucida.

DISCUSIÓN DE RESULTADOS
Los contaminantes atmosféricos pueden ser retenidos por los líquenes, en partículas atrapadas en los espacios intercelulares. La retención eficiente de elementos contaminantes por parte del liquen dependerá del número y naturaleza del sitio de unión extracelular, edad del tejido y las condiciones de crecimiento del liquen. A pesar del daño que sufren la integridad de sus células y tejidos por la acumulación de sustancias particulares, los líquenes pueden indicar la presencia de compuestos químicos en la atmósfera urbana (Giordano et al. 2005).
Se pudo observar que de acuerdo en los tipos de talos homoeros la distribución irregular de las hifas del hongo y los gonidios y Heteromeros en la cual las hifas y el gonidio se distribuyen de forma ordenada formando capas: en primer lugar, el córtex superior, formado por una masa de hifas muy apretada. Después, una capa gonidial, formada por la mezcla de hifas y gonidios, que son más numerosos. La tercera capa es medular, en la que predominan las hifas. La cuarta capa es el córtex inferior, que consiste en una masa de hifas muy apretadas.
CONCLUSIONES
-Esta práctica permitió el conocimiento integral de la morfología interna de los líquenes estudiados, haciendo una distinción específica de sus prototipos, entendiendo que las estructuras que conforman a un liquen dependen de su clase o tipo por lo que se hace innecesario reconocerlos a simple vista y relacionarlos
-Se lograron observar las diferentes estructuras de los líquenes que los diferencian de los demás donde algunos presentaban varias capas de diferentes colores y formas y poseían las esporas el lugares específicos según su tipo
-Se identificaron los diferentes tipos de líquenes por su forma, textura, estructuras, esporas, y su corte.
- Se logro realizar el 100% del laboratorio sellando las cuatro placas de líquenes fruticuloso, Gelatinoso, Folioso, Ascocarpo ademas de realizar su respectivo análisis, descripción y dibujo.
CONCLUSIONES
-Esta práctica permitió el conocimiento integral de la morfología interna de los líquenes estudiados, haciendo una distinción específica de sus prototipos, entendiendo que las estructuras que conforman a un liquen dependen de su clase o tipo por lo que se hace innecesario reconocerlos a simple vista y relacionarlos
-Se lograron observar las diferentes estructuras de los líquenes que los diferencian de los demás donde algunos presentaban varias capas de diferentes colores y formas y poseían las esporas el lugares específicos según su tipo
-Se identificaron los diferentes tipos de líquenes por su forma, textura, estructuras, esporas, y su corte.
- Se logro realizar el 100% del laboratorio sellando las cuatro placas de líquenes fruticuloso, Gelatinoso, Folioso, Ascocarpo ademas de realizar su respectivo análisis, descripción y dibujo.
BIBLIOGRAFÍA
-Barreno, E. and Rico, V. (2018). SOBRE LA BIOLOGÍA DE LOS LÍQUENES. I. ANATOMÍA, MORFOLOGÍA Y ESTRUCTURAS VEGETATIVAS. [online] Revistas.um.es. Available at: http://revistas.um.es/analesbio/article/view/70291/67761 [Accessed 29 Apr. 2018].
-nina (2018). Hongos liquenizados. [online] Slideshare.net. Available at: https://www.slideshare.net/997227832/hongos-liquenizados [Accessed 29 Apr. 2018].
-Lijteroff, R., Lima, L. and Prieri, B. (2018). Uso de líquenes como bioindicadores de contaminación atmosférica en la ciudad de San Luis, Argentina. [online] Scielo.org.mx. Available at: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0188-49992009000200006 [Accessed 29 Apr. 2018].



A mi padre le diagnosticaron la enfermedad de Alzheimer, esta enfermedad neuronal comenzó 25 años antes de que se revelara que se desencadenó en el momento en que comenzamos a comer azúcares, incluidos todos y cada uno de los carbohidratos que se descomponen en glicerol, y se le asignó el mejor tratamiento médico aprobado por los Estados Unidos. Estados, (FDA) ha estado tomando colinesterasa, mientras tanto para tratar su pérdida de memoria, confusión y problemas con su pensamiento y razonamiento. Solo para ayudar a retrasar la progresión y controlar los síntomas, ya que no existe cura, leí con interés. Una vez pensé que la enfermedad de Alzheimer no tenía cura. Bueno, es cierto con la medicina inglesa, pero no con la medicina herbal. La experiencia de mi papá me abrió los ojos a la realidad de una cura a través de hierbas. Mi padre era un hombre vibrante antes de su encuentro con el Alzheimer. Era un científico, por lo que su capacidad mental nunca estuvo en duda. En 2013, le diagnosticaron Alzheimer. Los síntomas se manifestaron mediante la repetición de conversaciones y el olvido gradual de las cosas. Pasó de encontrar las palabras adecuadas durante la conversación a una confusión significativa y al olvido de detalles sobre sí mismo. No fue una buena experiencia ver a su padre, cuya brillantez no tenía rival, se convirtió totalmente en una sombra de sí mismo. Sus médicos dijeron que la enfermedad no tiene cura, solo se le administraron medicamentos para el tratamiento que tenían muchos efectos secundarios. A principios de 2016, mientras estaba en Internet, me encontré con una historia de Alzheimer y leí sobre una cura a través de hierbas con interés. Investigué más para asegurarme de que no era un engaño. En mi búsqueda, en el testimonio se mencionó la mezcla de hierbas medicinales del Doctor James. Me comuniqué con el Gran Sanador en su correo electrónico que se proporcionó (DRJAMESHERBALMIX@GMAIL.COM) y obtuve su medicina de mezcla de hierbas para mi padre. Me dijo que su medicina de mezcla de hierbas ayudará a mi papá a reducir el pliegue de proteína anormal dentro de sus neuronas y regular los nutrientes y moléculas en el sistema de su cuerpo, y también detendrá el trastorno progresivo que se acumula al dañar las células cerebrales y ayuda a que sus células débiles que hacen que el cerebro se encoja funcione bien, es una buena bebida herbal para la reparación celular. Este doctor James es un gran hombre y su medicina de mezcla de hierbas es maravillosa y funciona de manera efectiva como él dijo, sin efectos secundarios. Han pasado 4 años y mi papá está perfectamente bien y ha vuelto a su trabajo de laboratorio incluso en la vejez. En el caso de sus seres queridos con Alzheimer o demencia, quítelos de la medicina inglesa y use la mezcla de hierbas del Dr. James para el tratamiento. Si padece demencia, enfermedad de Alzheimer o enfrenta diferentes tipos de desafíos de salud, no dude en comunicarse con el gran sanador en su correo electrónico DRJAMESHERBALMIX@GMAIL.COM y obtener su cura permanente. Es un buen hombre y te ayudará a curar enfermedades como. Enfermedades cardiovasculares, cáncer, diabetes, hepatitis A, B C, enfermedad de Alzheimer, vih / sida, virus del herpes genital, esquizofrenia, trastornos bipolares, fibromas,
ResponderEliminar